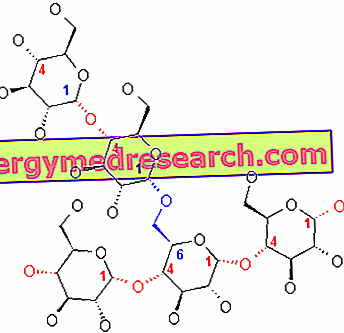
Glikogen adalah makromolekul (jisim molekul kira-kira 400 juta dalton) daripada α-glukosa di mana terdapat terutamanya ikatan-ikatan α-1, 4 glikosida dan cawangan dalam nisbah 1:10, disebabkan oleh ikatan-ikatan glikosida α-1, 6.
Glikogen ini merupakan bahan rizab dan terus direndahkan dan direkodkan semula; dalam semua jisim sel badan, kita mempunyai kira-kira 100 g glikogen: kebanyakannya adalah di dalam hati di mana ia adalah mudah alih dan boleh, oleh itu, digunakan sebagai rizab untuk organ-organ lain (glikogen dalam otot tidak bergerak).
Enzim-enzim yang memangkinkan degradasi dan sintesis glikogen, semuanya dalam sitoplasma, oleh itu satu sistem peraturan diperlukan yang menjadikan satu laluan tidak aktif apabila yang lain aktif: jika glukosa boleh didapati, kedua akan ditukar kepada glikogen (anabolisme) yang merupakan rizab, sebaliknya, jika glukosa diperlukan, maka glikogen dihina (katabolisme).
Enzim yang terlibat terutamanya dalam degradasi glikogen adalah fosforilasi glikogen ; enzim ini mampu memecahkan ikatan α-1, 4 glikosida dengan menggunakan sebagai ejen litik, ortofosfat anorganik: pembelahan berlaku melalui fosforikol dan glukosa 1-fosfat diperolehi.
Pada lima atau enam unit dari titik cawangan, enzim glikogen fosforilasi tidak lagi dapat bertindak, oleh itu ia melenyapkan dari glikogen dan digantikan oleh enzim debranching yang merupakan transferase : di tapak pemangkin enzim ini terdapat 'histidin yang membolehkan pemindahan tiga unit sakccharidic ke rantai glikosida terdekat (histidine menyerang karbon pertama dari molekul glukosa). Enzim yang baru disebutkan ialah glycosyltransferase ; pada akhir tindakan enzim ini, hanya satu unit glukosa kekal di rantaian sampingan dengan karbon pertama yang terikat pada karbon keenam glukosa dalam rantai utama. Unit glukosa terakhir rantaian sampingan dikeluarkan oleh tindakan enzim α-1, 6 glikosidase (enzim ini adalah bahagian kedua enzim debranching); memandangkan cawangan, dalam glikogen, berada dalam nisbah 1:10, dari degradasi lengkap makromolekul kita memperoleh kira-kira 90% glukosa 1-fosfat dan kira-kira 10% glukosa.
Tindakan enzim tersebut membolehkan penghapusan rantaian sampingan dari molekul glikogen; aktiviti enzim-enzim ini boleh diulang sehingga terdapat kemusnahan rantai yang lengkap.
Mari kita pertimbangkan hepatosit; glukosa (diasimilasikan melalui diet), apabila ia memasuki sel itu ia ditukar kepada glukosa 6-fosfat dan dengan itu diaktifkan. Glukosa 6-fosfat, dengan tindakan phosphoglucomutase, diubah menjadi glukosa 1-fosfat: yang terakhir adalah pendahulunya bukan biosintesis; dalam biosintesis, satu bentuk gula aktif digunakan, yang diwakili oleh gula yang terikat kepada dosis: biasanya uridyldiposfat (UDP). Glukosa 1-fosfat kemudian ditukar kepada UDP-glukosa; metabolit ini berada di bawah tindakan sintesis glikogen yang mampu mengikat UDP-glukosa ke akhir tanpa pengurangan glikogen yang semakin meningkat: glikogen yang dipanjangkan diperolehi dari unit glukosidik dan UDP. UDP ditukarkan oleh enzim nukleosidedifosphokinase ke UTP yang kembali ke peredaran.
Degradasi glikogen terjadi oleh tindakan fosforilase glikogen yang melepaskan molekul glukosa dan mengubahnya menjadi glukosa 1-fosfat. Selanjutnya, phosphoglucomutase menukarkan glukosa 1-fosfat kepada glukosa 6-fosfat.
Glikogen disintesis, di atas semua, di dalam hati dan di dalam otot: dalam badan terdapat 1-1.2 gram glikogen yang diedarkan di seluruh jisim otot.
Glikogen miokyte mewakili rizab tenaga hanya untuk sel itu manakala glikogen yang terkandung dalam hati juga merupakan rizab untuk tisu lain, iaitu, ia boleh dihantar, sebagai glukosa, ke sel lain.
Glukosa 6-fosfat yang diperolehi dalam otot-otot dari degradasi glikogen kemudiannya dihantar, dalam keadaan keperluan tenaga, kepada glikolisis; di dalam hati, glukosa 6-fosfat diubah menjadi glukosa oleh tindakan glukosa 6-fosfat fosfatase (satu ciri enzim hepatosit) dan disampaikan ke dalam aliran peredaran darah.
Kedua-dua fosforilase glikogen sintesis dan glikogen berfungsi pada unit glikogen yang tidak berkurang, jadi mesti ada isyarat hormon yang memerintahkan pengaktifan satu laluan dan menyekat yang lain (atau sebaliknya).
Di makmal, mungkin untuk memanjangkan rantai glikogen, mengeksploitasi fosforilasi glikogen dan menggunakan glukosa 1-fosfat dalam kepekatan yang sangat tinggi.
Di dalam sel, fosforilasi glikogen hanya memangkinkan tindak balas degradasi kerana kepekatan metabolit adalah seperti mengalihkan keseimbangan tindak balas berikut, ke kanan (iaitu ke arah kemerosotan glikogen):

Mari lihat pada mekanisme tindakan phosphorylase glikogen: oksigen asetik (yang berfungsi sebagai jambatan antara unit glukosa) mengikat kepada hidrogen fosforil: pertengahan tindakbalas terbentuk oleh karbokatione (pada glukosa yang hujung) di mana fosforil (Pi) mengikat dengan cepat.
Phosphorylase glycogen memerlukan cofactor yang fosfat pyridoxal (molekul ini juga merupakan cofactor untuk transaminase): ia mempunyai fosforil protonated hanya sebahagian (fosfat pyridoxal dikelilingi oleh persekitaran hidrofobik yang membenarkan kehadiran proton terikat kepadanya) . Fosforil (Pi) dapat menghasilkan proton kepada glikogen kerana reaksi fosforil tersebut kemudian, proton dari fosforil sebahagian proton fosfat pyridoxal. Kebarangkalian bahawa, pada pH fisiologi, fosforil kehilangan proton dan masih sangat deprotonated sangat rendah.
Sekarang mari kita lihat bagaimana phosphoglucomutase berfungsi. Enzim ini mempunyai residu serum fosforilasi di tapak pemangkin; serina menghasilkan fosforil ke glukosa 1-fosfat (dalam kedudukan enam): glukosa 1, 6-bisfosfat terbentuk untuk masa yang singkat, maka serine akan diofosilasi dengan mengambil fosforil dalam kedudukan satu. Phosphoglucus mutase boleh berfungsi di kedua-dua arah, iaitu mengubah glukosa 1-fosfat ke glukosa 6-fosfat atau sebaliknya; jika glukosa 6-fosfat dihasilkan, ia boleh terus dihantar ke glikolisis, ke dalam otot, atau diubah menjadi glukosa dalam hati.
Enzim transfer uridyl phosphoglucus (atau UDP glukosa pyrophosphorylase) memangkinkan tindak balas pemindahan glukosa 1-fosfat kepada UTP oleh serangan phosphoryl a.
Enzim yang diperihalkan adalah pyrophosphorylase: nama ini disebabkan oleh kenyataan bahawa tindak balas yang bertentangan dengan yang baru diperihalkan adalah pyrophosphosphorylation.
Glukosa UDP, yang diperolehi seperti yang dijelaskan, mampu memanjangkan rantai glikogen unit monosakarida.
Ia boleh mengubah reaksi terhadap pembentukan glukosa UDP dengan menghapuskan produk yang pirofosfat; pyrophosphatase enzim menukarkan pirofosfat ke dalam dua molekul ortofosphat (hidrolisis anhidrida) dan dengan demikian, mengekalkan kepekatan pyrophosphate begitu rendah sehingga proses pembentukan glukosa UDP disukai termodinamik.
Seperti yang dinyatakan, glukosa UDP, terima kasih kepada tindakan sintesis glikogen, dapat memanjangkan rantai glikogen.
Pengaruh (dalam nisbah 1:10) disebabkan oleh fakta bahawa, apabila rantai glikogen terdiri daripada 20-25 unit, enzim bercabang (mempunyai histidin di tapak pemangkinnya) yang campur tangan, mampu memindahkan satu siri 7 -8 unit glikosidik lebih jauh di hilir 5-6 unit: cawangan baru dihasilkan.
Atas alasan asal saraf atau jika tenaga diperlukan kerana usaha fizikal, adrenalin disembur dari kelenjar adrenal.
Sel sasaran adrenalin (dan norepinefrin) adalah hati, otot dan tisu adipose (di dalamnya terdapat penurunan trigliserida dan peredaran asid lemak: dalam mitokondria, oleh itu, menghasilkan glukosa 6 -fosfat, dihantar ke glikolisis, manakala dalam adiposit, glukosa 6-fosfat diubah menjadi glukosa oleh tindakan enzim glukosa 6-fosfat fosfatase dan dieksport ke tisu).
Mari lihat, sekarang, mod tindakan adrenalin. Adrenalin akan mengikat kepada reseptor yang diletakkan pada membran sel (daripada myocytes dan hepatosit) dan ini menentukan terjemahan isyarat dari luar ke dalam sel. Kinase protein diaktifkan yang bertindak serentak pada sistem yang mengawal sintesis dan degradasi glikogen:
Synthase glikogen wujud dalam dua bentuk: bentuk dephosphorylated (aktif) dan bentuk phosphorylated (tidak aktif); phosphorylates protein kinase glycogen synthase dan blok tindakannya.
Fosforilasi glikogen boleh wujud dalam dua bentuk: satu bentuk aktif di mana serina fosforilasi hadir dan suatu bentuk tidak aktif di mana serina dilepaskan. Fosforilasi glikogen boleh diaktifkan oleh kinase fosforilase glikogen enzim. Kinase phosphorylase glikogen aktif jika ia fosforilasi dan tidak aktif jika ia dilepaskan; kinase protein mempunyai substrat kinase fosforilase glikogen, iaitu ia dapat memfosforat (dan oleh itu, mengaktifkan) yang kemudiannya mengaktifkan fosforilasi glikogen.
Apabila isyarat adrenalin selesai, kesannya dalam sel mesti berakhir: enzim fosfatase campur tangan, kemudian, pada spesies protein.